|
Fondazione FONAMA |
fonama@fonama.org |
Tel. 335250742 |
 |
 |
Traduzioni a cura di Natale Marzari
Dopo 41 anni e 5 mesi, nel maggio 2006 la
magistratura di Trento ha riconosciuto l'esistenza e la gravità di quella
malattia rara che nessuna altra istituzione o persona singola della provincia di
Trento ancora mi riconosce, e per negare la quale ancora mi perseguita.
Natale Marzari |
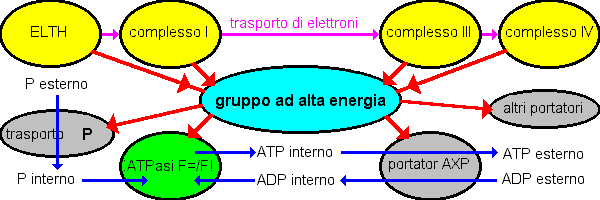
Il sistema ad alta energia.
Esperimenti di trasporto
inverso di elettroni portarono negli anni 1950 a realizzare che dovrebbe esserci un comune intermedio non-fosforilato ad alta energia tra
i portatori della catena respiratoria e la sintesi dell' ATP. Questo concetto è illustrato sotto. Le frecce mostrano la direzione normale
del flusso, ma la maggior parte delle reazioni sono liberamente reversibili, e l'energia aggiunta al sistema via un processo può essere utilizzata da qualunque altra parte. Il
sistema ad alta energia fu originalmente pensato essere una combinazione chimica, ma oggigiorno noi sappiamo che esso è in realtà il pH ed i gradienti elettrici sviluppati attraverso la
membrana interna mitocondriale.
E' possibile misurare i gradienti studiando la distribuzione di molecole permeanti attraverso la membrana mitocondriale interna. Questo richiede un metodo per la rapida separazione dei
mitocondri dal loro mezzo di incubazione, sia attraverso una rapida ultrafiltrazione, o
con una centrifugazione attraverso un strato di olio di silicone. Un marcatore impermeabile
radioattivo (comunemente saccarosio) viene aggiunto alla incubazione al fine di corregge il mezzo esterno contaminante gli organelli.
Gli acidi grassi debolmente solubili possono essere usati per misurare il differenziale pH.
Questo metodo sfrutta la facile permeabilità della membrana agli acidi liberi
elettricamente neutri, e l'impermeabilità ai loro relativi sali ionizzati.
È vitale che l'acido libero attraversi la membrana in forma protonata senza
movimento netto di cariche elettriche. In equilibrio la concentrazione
dell'acido libero è la stessa dappertutto, ma il rapporto fra [ acido libero ] e
[ sale ionizzato ] differisce nei due scomparti. Nello spazio della matrice la
quantità di sale è proporzionale al pH interno, ma fuori dei mitocondri la
concentrazione ionizzata nel sale dipende dal pH esterno. Se le concentrazioni
interne ed esterne del sale possono essere
misurate allora l'applicazione dell'equazione
di Henderson Hasselbach ci rende noto il pH interno.
La misura del potenziale di membrana ( )
richiede un catione permeante, per es. un metallo alcalino radioattivo più valinomicina,
o il tetrafenilfosfonio solubile nei del lipidi. In questo caso è importante che
il catione penetri la membrana con netto movimento di carica, in modo che
la distribuzione del catione sia influenzata dal potenziale di membrana. Se le
concentrazioni interne ed esterne di un catione permeant possono essere
misurate, allora il potenziale della membrana può essere calcolato
dall'equazione del equazione di Nernst.
)
richiede un catione permeante, per es. un metallo alcalino radioattivo più valinomicina,
o il tetrafenilfosfonio solubile nei del lipidi. In questo caso è importante che
il catione penetri la membrana con netto movimento di carica, in modo che
la distribuzione del catione sia influenzata dal potenziale di membrana. Se le
concentrazioni interne ed esterne di un catione permeant possono essere
misurate, allora il potenziale della membrana può essere calcolato
dall'equazione del equazione di Nernst.
Alcuni coloranti cambiano le loro proprietà di fluorescenza o di assorbimento
quando sono stati allineati in un campo elettrico forte. Ciò fornisce un metodo
alternativo per la misura continua del potenziale della membrana mitocondriale
aggiungendo tali sostanze ad una sospensione mitocondriale. Le sostanze adatte
includono le carbocianine, le fenosafranine ed i bisossonoli. Anche se è molto
conveniente, questa tecnica deve essere calibrata usando il più vecchio metodo
del punto singolo.
La somma del potenziale della membrana ed il gradiente pH sono conosciute
insieme come energia motrice del protone
(PMF). Ciò indica l'energia potenziale totale immagazzinata nel gradiente
transmembranico, il quale è a disposizione per guidare i protoni nuovamente
dentro la matrice , e fornire l'energia per i processi biologicamente utili.
Usando il segno corretto per il gradiente pH (negativo!) e sostituendo i valori
numerici per la costante di gas, la costante di Faraday e la temperatura
assoluta, ne segue questo
PMF (in millivolt) = -
60 pH
pH
a 37oC. Il segno minus è confondente: il gradiente pH è egli stesso negativo,
così i due componenti sono normalmente cumulativi nei loro effetti.
Nei mitocondri il gradiente elettrico,(150mV,
interno negativo) dà un contributo più grande che il gradiente pH (0.5pH unità ,
interno alcalino). Nei cloroplasti il pH
è più importante. Il pH
mitocondriale corrisponde ad una differenza tripla nella concentrazione dello
ione di idrogeno fra la matrice ed il citosol. Per ogni processo del
transmembranico, il
pH ed il
componenti possono agire sia separatamente sia insieme, secondo la struttura
degli enzimi e l'equilibrio del vantaggio biologico. Un movimento del protone
trasporterebbe normalmente una carica positiva, ma possono muoversi pure altri
ioni, così non c'è esigenza del protone ed i conteggi della carica per
equilibrare per ogni individuale processo di trasporto membranale, anche se i
totali devono equilibrare ovviamente globalmente.
Le migliori valutazioni correnti migliori per i conteggi della carica e del
protone per la catena respiratoria sono indicate nella sezione sugli elementi
portatori redox. Per ogni paio di
elettroni che attraversano la catena, La NADH deidrogenasi sposta 4 protoni e 4
cariche positive esternamente attraverso la membrana interna dalla matrice verso
il citosol, la citocromo c riduttasi dell'ubichinolo sposta 4 protoni e 2
cariche, mentre la citocromo c ossidasi sposta 2 protoni e 4 cariche. La
ri-ossidazione del NADH per formare il NAD e l'acqua è così associata con
l'esportazione netta di 10 protoni e di 10 cariche positve dalla matrice
mitocondriale al compartimento citosolico.
La sintesi dell'ATP dalla F1ATPasi si pensa richiedere 3 protoni che
trasportano 3 cariche positive per ri-entrare nella matrice. In più, 1 protone
(ma nessuna carica) è necessari per l'assorbimento del fosfato e 1 carica (ma
nessun protone) per lo scambio ATP/ADP con il citosol (vedi le pagine
trasporto dei metaboliti per i particolari).
Così il requisito generale è 4 protoni e 4 cariche per ogni ATP trasportato al
mondo esterno. Ciò implica che il rapporto P:O è 2,5 per i substrati
NAD-collegati, e 1,5 per il succinato, il quale è un poco più basso dei valori
riportati nei manuali più vecchi, ma molto più vicino ai risultati sperimentali
reali!
Il doppio strato lipidico della membrana interna è
spesso soltanto circa 5nm ed è quindi conforme alle forze elettriche enormi:
150 x 10-3 volt per 5 x 10-9 metri
= 30,000,000 volt / metro
La membrana interna ha una composizione insolita in fosfolipidi
che include il
cardiolipin (difosfatodilglicerolo) ed è specialmente
"prova della perdita" di piccoli ioni.
The inner membrane has an unusual phospholipid composition which includes
cardiolipin (diphosphatidylglycerol) and is particularly
"leak proof" to small ions.
La maggior parte dei tipi di fosfolipidi formeranno spontaneamente i liposom
una volta agitati con acqua, e questo è stato sfruttato per ri-costruire
la catena respiratoria dai componenti purificati.