|
Fondazione FONAMA |
fonama@fonama.org |
Tel. 335250742 |
 |
 |
Traduzioni a cura di Natale Marzari
Dopo 41 anni e 5 mesi, nel maggio 2006 la
magistratura di Trento ha riconosciuto l'esistenza e la gravità di quella
malattia rara che nessuna altra istituzione o persona singola della provincia di
Trento ancora mi riconosce, e per negare la quale ancora mi perseguita.
Natale Marzari |
Breve storia e orientamento
Gli spettri dei pigmenti respiratori del citocromo nei tessuti
animali sono stati osservati originalmente nel diciannovesimo secolo, ma
allontanati come manufatto dall'istituzione scientifica del tempo. Il consumo
d'ossigeno dagli omogenati di tessuti, e l'azione degli inibitori respiratori
semplici in primo luogo è
stata studiata sistematicamente negli anni 1930, quando è stato realizzato che gli
elettroni scorrono dai
substrati ossigeno via una sequenza di elementi portatori riducenti, molti dei
quali
mostrano cambiamenti spettrali distinti se
ossidati o ridotti.
L'isolamento dei mitocondri intatti ha seguito i miglioramenti nella
tecnologia della centrifuga negli anni 1940, e l'introduzione dei mezzi isotonici del
preparato di il saccarosio
per impedire la lisi osmotica. I ricercatori hanno notato il fenomeno della latenza: gli enzimi mitocondriali interni non poterono essere dimostrati fino
a che gli organelli non furono rotti ed aperti. La struttura di base
dei mitocondri è stata stabilita con la microscopia
elettronica ed è stata rivelata tramite rottura controllata della
maggior parte degli enzimi respiratori responsabili del consumo
d'ossigeno fissati alla membrana mitocondriale interna
(la quale è piegata estesamente per formare le creste mitocondriali)
mentre gli enzimi solubili che catalizzano le vie ossidative del
ciclo di Krebs e degli acidi grassi sono confinati nella
matrice interna.
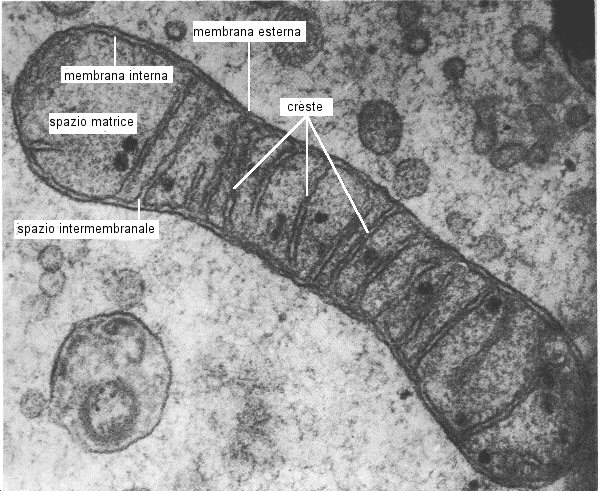 |
Questa
è un'immagine al microscopio elettronico di un mitocondrio
dell'embrione di pulcino ed è usata con il gentile permesso del professor
Ruth Bellairs, del Reparto di anatomia e di biologia inerente allo sviluppo,
Università Dell'Università, Via Di Gower, Londra WC1E 6BT.
|
Più tardi veniva scoperto che la membrana interna era una barriera
importante alla permeabilità all'interno della cellula, ma la
membrana esterna
contiene una proteina denominata porina che
la rende in gran parte permeabile alle molecole minori di circa
1500 dalton. Un gruppo importante di enzimi che metabolizzano l'ATP, comprende
la
miochinasi, la creatina chinasi, e la
nucleoside disolfato chinasi, sono
confinate nello spazio inter-membranale, fra
le membrane interne ed esterna.
miochinasi: ATP + AMP <=> ADP + ADP
creatina chinasi: ATP + creatina <=> ADP + creatine fosfato
citidine difosfato chinasi: CDP + ATP <=> CTP + ADP (esistono molti
enzimi simili)
Il lavoro sperimentale è accelerato con l'introduzione dell'elettrodo ad
ossigeno Clarke-tipo più conveniente negli anni '50 al
posto dei più vecchi metodi manometrici. È stato indicato che l'assorbimento di
ossigeno dai mitocondri
intatti dipendeva dalla conversione simultanea dell'ADP e del fosfato
inorganico in ATP. Il fenomeno è chiamato controllo
respiratorio. La dipendenza obbligatoria di una reazione chimica altamente
favorevole (ossidazione del substrato) sull'esecuzione simultanea di
una reazione indipendente ed estremamente sfavorevole (sintesi dell'ATP) è stata riconosciuta
immediatamente come un
nuovo e importante concetto dai biochimici di allora
.
Il consumo d'ossigeno che dipende dalla presenza dell'ADP e del
fosfato è chiamata respirazione accoppiata. L'accoppiamento della respirazione e
della fosforilazione ha potuto
essere rotto tramite rottura meccanica o osmotica dei mitocondri, o tramite
l'aggiunta di agenti disaccoppianti, nel qual caso la respirazione continua
velocemente senza alcuna
sintesi concomitante di ATP. L'agente disaccoppiante
originale era il dinitrofenolo 2,4, il quale ora è stato sostituito da sostanza più
efficaci quali il FCCP
(fenilidrazone del cianuro del carbonilico di
paragrafi-trifluorometossile) e CCCP (fenilidrazone meta-cloro del
cianuro carbonilico).
Le due maggiori categorie di inibitori mitocondriali
possono essere distinte in: inibitori della respirazione ed inibitori della fosforilazione. Entrambi i tipi
sono efficaci contro i mitocondri intatti, ma
soltanto gli inibitori della respirazione hanno potuto impedire il
consumo d'ossigeno dopo l'aggiunta di un agente disaccoppiante. Questi esperimenti
hanno rafforzato il concetto di un sistema energico
favorevole di trasporto dell'elettrone, il quale in qualche modo alimentata un
sistema energico sfavorevole di
fosforilazione. L'energia libera disponibile dalle reazioni di riduzione è stata usata
per
guidare la sintesi dell'ATP. Gli inibitori della respirazione
quali gli ioni del cianuro
ostruiscono la catena di trasporto dell'elettrone ed erano sempre
efficaci nella soppressione del consumo d'ossigeno, mentre gli inibitori
della fosforilazione quale la il oligomicina,
che impediscono la fabbricazione dell'ATP, possono bloccare la respirazione soltanto se
l'accoppiamento fosse
intatto.
A questo punto, alla metà dagli anni 50, ci si trovava in un vicolo cieco lungo
ed occasionalmente
acrimonioso, dove gli scienziati tentavano di trovare un mediatore di
collegamento chimico fra la respirazione e la generazione di ATP. Questa idea derivava dai
livelli di fosforilazione del
substrato
osservati nella via glicolitica. Questa ricerca era infruttuosa poiché non
esiste nessuno di
tali intermediarie. Ora sappiamo che l'energia proveniente dalla respirazione in primo luogo
è bloccata sotto forma di pH ed i gradienti dei potenziali elettrici
attraverso la membrana mitocondriale interna. L'energia immagazzinata in
questi gradienti è successivamente
sfruttata per guidare la sintesi dell'ATP. Questa
teoria chemiosmotica è stata proposta in primo luogo nel 1961 da
un biochimico britannico, Peter
Mitchell,, che ha lavorato per molti anni senza
molto riconoscimento ufficiale o
fondi pubblici significativi fino a che non ricevette
il premio Nobel per la chimica nel 1978.
{kind=link}