|
Fondazione FONAMA |
fonama@fonama.org |
Tel. 335250742 |
 |
 |
Traduzioni a cura di Natale Marzari
Dopo 41 anni e 5 mesi, nel maggio 2006 la
magistratura di Trento ha riconosciuto l'esistenza e la gravità di quella
malattia rara che nessuna altra istituzione o persona singola della provincia di
Trento ancora mi riconosce, e per negare la quale ancora mi perseguita.
Natale Marzari |
Ossigrafia - misurazione con l'elettrodo ad ossigeno
Quando i mitocondri sono incubati in un
apparato ossigrafico (elettrodo ad ossigeno) in un mezzo isotonico contenente substrato
e fosfato, l'aggiunta di ADP causa un improvviso picco di consumo di ossigeno
quando l'ADP viene convertito in ATP:
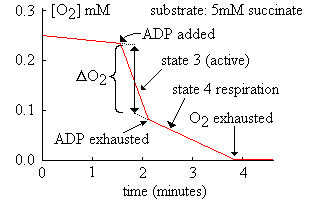
L'attività respiratoria rapida è chiamata "stato
3" della respirazione, mentre il tasso respiratorio più lento dopo che tutta l'ADP è stata fosforilata
per formare ATP è chiamato "stato 4".
Lo stato respiratorio 4 è solitamente più rapido della
velocità originale prima
della prima aggiunta di ADP poiché alcune ATP vengono demolite dalla attività
della ATPasi contaminante il preparato, e l'ADP derivante viene poi ri-fosforilata
nei mitocondri intatti.
Il rapporto fra [velocità stato 3] : [velocità stato 4] è chiamato
indice di controllo respiratorio (RCI) e indica la strettezza dell'accoppiamento
fra la respirazione e la fosforilazione. Nei mitocondri isolati l'accoppiamento
non è perfetto, probabilmente come risultato del danneggiamento meccanico durante
le procedure di isolazione. I valori tipici del RCI variano in una gamma da 3 a 10, varianti
con il substrato e la qualità della preparazione. Si pensa
che l'accoppiamento sia migliore in vivo, ma non può però arrivare al 100%.
E' possibile calcolare un rapporto
P:O (il rapporto fra la sintesi di ATP ed il consumo di ossigeno) misurando
la diminuzione del tenore dell'ossigeno durante il rapido picco dello stato 3
della respirazione dopo aver aggiunto un quantitativo conosciuto di ADP. E' necessario sottrarre
la respirazione basale dovuta all'accoppiamento imperfetto ed al riciclaggio dell'ATP,
come mostrato nel grafico sopra. Il cambiamento nella concentrazione deve essere
quindi moltiplicato
per il volume della camera, in modo che la risposta (in micro-atomi di ossigeno)
possa essere correlata alla quantità di ADP aggiunta. La quantità di ossigeno nella camera
viene calcolata dai dati pubblicati della solubilità dell'ossigeno alla
temperatura adatta. Usando la figura del grafico qui sopra:
diminuzione in O2 (dal grafico) = 0,135 mM; volume della camera =
2,5ml
atomi di ossigeno consumati nello stato 3 = 0,135 * 2 * 2,5 = 0,68 micro-atomi
(nota il fattore di 2, tenendo conto di 2 atomi in una molecola di ossigeno)
iniettato ADP (20 microlitri di una soluzione a 50mM) = ATP formato = 1 micromole
in totale
P:O = [ATP formato] : [ossigeno consumato] = 1,48 (per ossidazione del succinato)
I substrati collegati a NAD danno valori consistentemente più alti per
il rapporto P:O
(circa 2,5) paragonati con il succinato (circa 1,5). Questi risultati indicano
che gli elettroni dagli agenti relativamente
scarsamente riducenti tipo il succinato (anche l'acil CoA ed il glicerolo fosfato) entrano
a parte lungo il percorso della catena respiratoria, saltando il primo sito di accoppiamento
dove l'energia è catturata per la sintesi dell'ATP.
Le variazioni di forma della linea inclinata dopo l'esaurimento dell'ADP e nuovamente dopo
che tutto
l'ossigeno è stato usato implicano che i mitocondri devono avere una affinità molto alta per
l'ADP e per l'ossigeno. La concentrazione di entrambe queste sostanze
è molto bassa nella maggior parte delle cellule sane, poiché esse ne sono
efficentemente ripulite dai mitocondri.
L'aggiunta di un agente di disaccoppiamento (come il dinitrofenolo o CCCP)
porta ad un tasso di respirazione permanentemente alto in assenza di ADP,
fino a che tutto l'ossigeno non è stato consumato.
Molti veleni naturali e sintetici bloccano la respirazione
mitocondriale.
Se i mitocondri sono incubati in un ossigrafo da esperimento con substrati
e fosfato inorganico, le interazioni fra inibitori
e disaccoppiatori permettono di
distinguere i due maggiore tipi
di inibizione:
| tassi respiratori mitocondriali |
|
|
|
| aggiunti / inibitori |
nessuno |
oligomicina |
cianuro |
| nessuno |
basso |
basso |
zero |
| ADP |
alto |
basso |
zero |
| agenti di disaccoppiamento |
alto |
alto |
zero |
La conclusione da trarre da questo tipo di esperimento è
che il cianuro
impedisce la respirazione bloccando la stessa catena respiratoria, così che il cianuro
è efficace sia che siano o non siano aggiunti ADP o un disaccoppiatore. L'oligomicina
d'altra parte inibisce il sistema di fosforilazione dell'ADP, così blocca solo
la
respirazione accoppiata nei mitocondri.