|
Fondazione FONAMA |
fonama@fonama.org |
Tel. 335250742 |
 |
 |
Traduzioni a cura di Natale Marzari
Dopo 41 anni e 5 mesi, nel maggio 2006 la
magistratura di Trento ha riconosciuto l'esistenza e la gravità di quella
malattia rara che nessuna altra istituzione o persona singola della provincia di
Trento ancora mi riconosce, e per negare la quale ancora mi perseguita.
Natale Marzari |
Strutture chimiche
Indice
Non dovete realmente impararli! Se dovete conoscere una struttura allora
osservatela. A volte aiuta a conoscere di quale genere di materiali state
occupandovi, e per chi lo vuole fare, o per coloro che sono più curiosi del normale
ho raccolto qui sotto le
strutture di alcune materie chiave che sono state usate per studiare
il metabolismo mitocondriale.
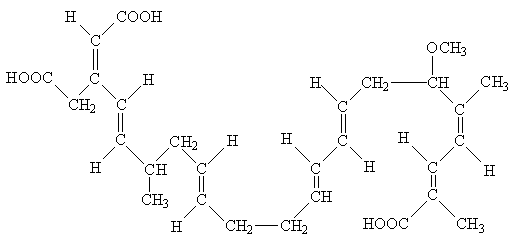
Acido bongcrecico: Questo inibitore tossico del portatore nucleotide adenina
è
ottenuto da una muffa indesiderata che a volte si sviluppa sul "bongkrek". Questo
piatto indonesiano fermentato sembra essere un ibrido fra
formaggio blu danese ed il budino di riso. L'inibitore si lega al portatore
nella conformazione "rivestimenti
interni", quando ha trasportato appena un carico fresco di ADP nella
matrice
mitocondriale. Di conseguenza, i mitocondri bongkrekate-inibiti
contengono sensibilmente più
nucleotidi degli organelli atrattilosido-inibiti. Ciò può essere usata
per valutare il loro contenuto molare di
proteine del portatore del nucleotide adenina.
Amino-ossiacetato: Questo derivato dell'idrossilammina reagisce chimicamente
con il
fosfato del piridossale e blocca tutte le reazioni delle
transaminasi.
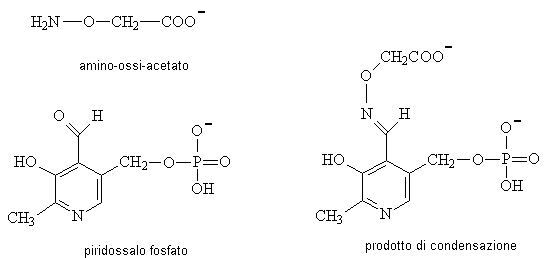
Inibisce il ciclo malato - aspartato, e così può essere usato valutare il
flusso attraverso
la glutammato deidrogenasi senza interferenze dalla via più
attiva di transaminazione.
Antimicina A: Questo potente inibitore potente del complesso 3
agisce fra il
citocromo
b ed il citocromo c1
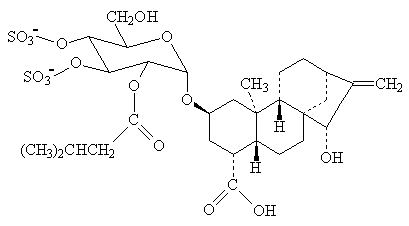
Atrattilosido: Questo inibitore tossico del portatore
adenina nucleotide
è
ottenuto dal cardo mediterraneo, atractylis gummifera. Si lega al portatore
nella conformazione "rivestimenti esterni", quando ha appena trasferito il
suo ATP nel
citosol e sta per legare un ADP per trasportarlo nella matrice mitocondriale.
Citocromi: Tutti i citocromi sono proteine eme, ma ci sono
leggere differenze
chimiche fra gli eme a, b e c con la conseguenza che le loro bande di
assorbimento visibili
sono spostate un po'. Si noti che queste sostanze subiscono l'ossidazione e la
riduzione virando, ciclando fra le forme ferrose e ferriche, contrariamente ad emoglobina
in cui il ferro è normalmente nello stato
ferroso. Il citocromo c (kd 12) è fra il più piccolo di queste molecole
e opera nello spazio intermembranico come portatore mobile di
riduttori equivalenti fra il complesso 3 ed il complesso IV.
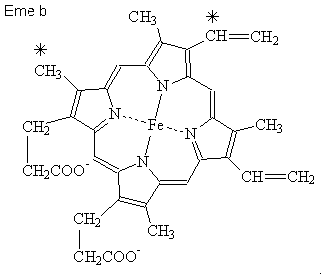 |
L'eme a assomiglia all'eme b salvo che il gruppo metilico
iniziale è
ossidato ad un gruppo formile, ed il gruppo iniziale del vinile è sostituito
da una catena laterale
di isoprenoide a 17-atomi di carbonio, più corto ma al contrario molto simile alla catena
laterale idrofoba
ramificata in Coenzyme
Q.
L'eme c è inoltre simile all'eme b, ma in questo caso entrambi i gruppi del vinile
dell'eme reagiscono con
i residui della cisteina nella proteina, in covalenza fissa il residuo eme alla catena del polipeptide.
|
Ferro non-eme: Questa importante categoria
di proteine è distribuita molto
ampiamente nelle piante, negli animali e nei microorganismi, dove sono impiegati in un'ampia
varietà di reazioni di riduzione. Ci ne sono centinaia dei tipi differenti, i quali
differiscono considerevolmente nelle loro strutture e nelle proprietà di riduzione, ma tutti contengono uno o più atomi del ferro circondati
dallo zolfo
- contenente l'aminoacido cisteina. Per questo motivo spesso sono chiamati
"proteine ferro - zolfo".
Flavine: Il derivati FAD e FMN della flavina sono sintetizzati da
riboflavina dietetica (vitamina B2). Si incontrano più comunemente come
gruppi prostetici, permanente fissati agli enzimi addetti alle reazioni redox,
dove funzionano come elementi portanti provvisori di riduzione degli
equivalenti come componente del meccanismo catalitico.
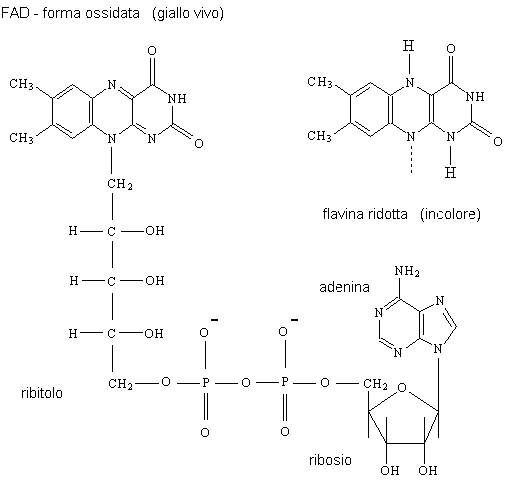
FMN è molto simile nelle proprietà al FAD, ma difettando della AMP
contiene soltanto la flavina, il ribitol (un alcool di zucchero derivato dal
ribosio) e fosfato. Entrambi i coenzimi sono il colore giallo luminoso nella
forma
ossidata, ma diventa slavato in riduzione.
Oligomicina:Questa sostanza inibisce la
sintesi dell'ATP (F0/F1 Atpasi) legando "il peptide conferitore di sensibilità
all'oligomicina" (OSCP) nella regione del gambo dell'enzima.
Piridina nucleotide: I coenzimi della piridina
nucleotideNAHD/ Nad e
NADPH/ Il NADP sono sintetizzato da nicotinammide (niacina, la vitamina B3) e
sono i principali portantori mobili di
riduttori equivalenti fra gli enzimi solubili della deidrogenasi
e la catena respiratoria.
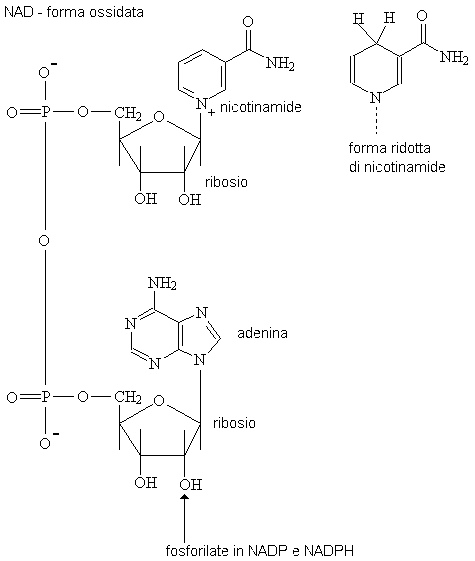
Le forme ridotte di entrambi i coenzimi assorbono fortemente la luce
ultravioletta intorno alla lunghezza d'onda di 340nm e una soluzione
di 0.1mM ha una densità ottica di 0.62 in una provetta chiara 1cm di
lunghezza del percorso (coefficente di estinzione = 6220 litritalpacentimetro).
Ciò costituisce la base per i metodi numerosi di analisi dell'enzima. In più, se
i coenzimi ridotti sono illuminati intorno a 340nm diventano
flourescenti fortemente intorno 450nm e le frazioni di una nanomole
sono immediastamente rilevate con questo metodo. È possibile controllare
l'ossidazione e la riduzione dei nucleotidi
della piridina di mitocondri isolati o persino di tessuti intatti
usando metodi ottici.
Rotenone: Questo inibitore potente del complesso mitocondriale
I è un
insetticida naturale derivato dalla radice di rotenone.
TTFA: TTFA: Questo inibitore del complesso 2 è usato nei processi metallurgici.
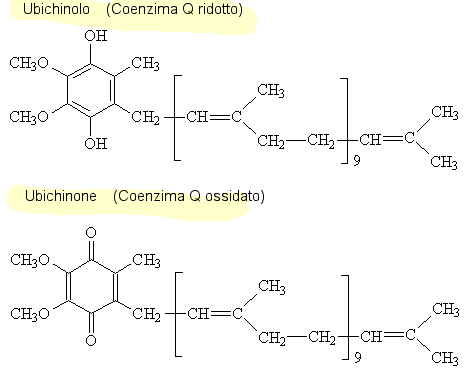
Ubichinolo / ubichinone: Come il suo nome suggerisce, questa
sostanza è distribuita molto
ampiamente in natura. Ci sono differenze di specie nella lunghezza
della catena laterale dell'isoprenoide. Tutte le forme naturali sono insolubili in acqua, ma
è una sostanza
solubile nei lipidi della membrana in cui funzionano come
portatore mobile di riduttori equivalenti fra i
complessi del multi-enzima.
Strutture chimiche